Nội dung chính
Vô sinh là một vấn đề sức khỏe sinh sản toàn cầu, ảnh hưởng khoảng 10 –15% các cặp vợ chồng trong độ tuổi sinh sản. Tỷ lệ vô sinh nữ đang có xu hướng gia tăng do nhiều nguyên nhân như suy buồng trứng sớm (Premature Ovarian Insufficiency - POI), suy giảm dự trữ buồng trứng theo tuổi và vô sinh liên quan đến điều trị ung thư (Cancer Treatment Related Infertility - CTRI). Trong đó, POI tác động đến khoảng 1% phụ nữ dưới 40 tuổi, trong khi các liệu pháp hóa trị hoặc xạ trị có thể gây ra những tổn thương không hồi phục đối với mô buồng trứng dẫn đến cạn kiệt nguồn noãn bào. [1, 2]
Mặc dù các kỹ thuật hỗ trợ sinh sản đã cải thiện đáng kể khả năng điều trị vô sinh, hiệu quả của các phương pháp này vẫn phụ thuộc chủ yếu vào số lượng và chất lượng noãn tự thân của bệnh nhân. Đối với những phụ nữ đã suy giảm hoặc mất hoàn toàn nguồn dự trữ buồng trứng, việc thu nhận noãn cho mục đích thụ tinh trong ống nghiệm thường gặp nhiều khó khăn hoặc không thể thực hiện được, buộc họ phải tìm đến phương án sử dụng noãn hiến tặng như lựa chọn thay thế cuối cùng.
Trong bối cảnh đó, sự phát triển của các mô hình biệt hoá noãn bào invitro từ tế bào gốc đã mở ra một hướng đi đột phá trong lĩnh vực sinh học sinh sản. Tế bào gốc là những tế bào nguyên bản đa tiềm năng có thể phân chia thành nhiều loại tế bào khác nhau để sửa chữa, phát triển và tái tạo. Tế bào gốc thường đến từ hai nguồn: tế bào phôi sớm và các mô trưởng thành. Các nghiên cứu gần đây cho thấy tế bào gốc vạn năng có khả năng biệt hóa thành tế bào giống tế bào mầm nguyên thủy (Primordial Germ Cell-like Cells -PGCLCs) và tiếp tục hình thành noãn bào chức năng trong điều kiện nuôi cấy mô phỏng vi môi trường buồng trứng, tạo tiền đề cho các ứng dụng trong điều trị vô sinh và nghiên cứu sinh học sinh sản. Các loại tế bào gốc chính đang được tập trung nghiên cứu bao gồm: tế bào gốc vạn năng (Pluripotent Stem Cells - PSCs), tế bào gốc buồng trứng (Ovarian Stem Cells - OSCs) và tế bào gốc trung mô (Mesenchymal Stem Cells - MSCs). [1, 3]
Bài tổng quan nhằm hệ thống hóa các tiến bộ mới nhất trong nghiên cứu tạo noãn bào in vitro từ các nguồn tế bào gốc khác nhau, tập trung phân tích cơ chế biệt hóa, và hiệu quả của từng loại tế bào gốc trong việc tái tạo nguồn noãn bào. Đồng thời, thảo luận về những thách thức kỹ thuật, rào cản đạo đức và triển vọng ứng dụng lâm sàng của công nghệ này trong việc giải quyết tình trạng suy giảm dự trữ buồng trứng trong tương lai.
- Quá trình phát triển của nang noãn:
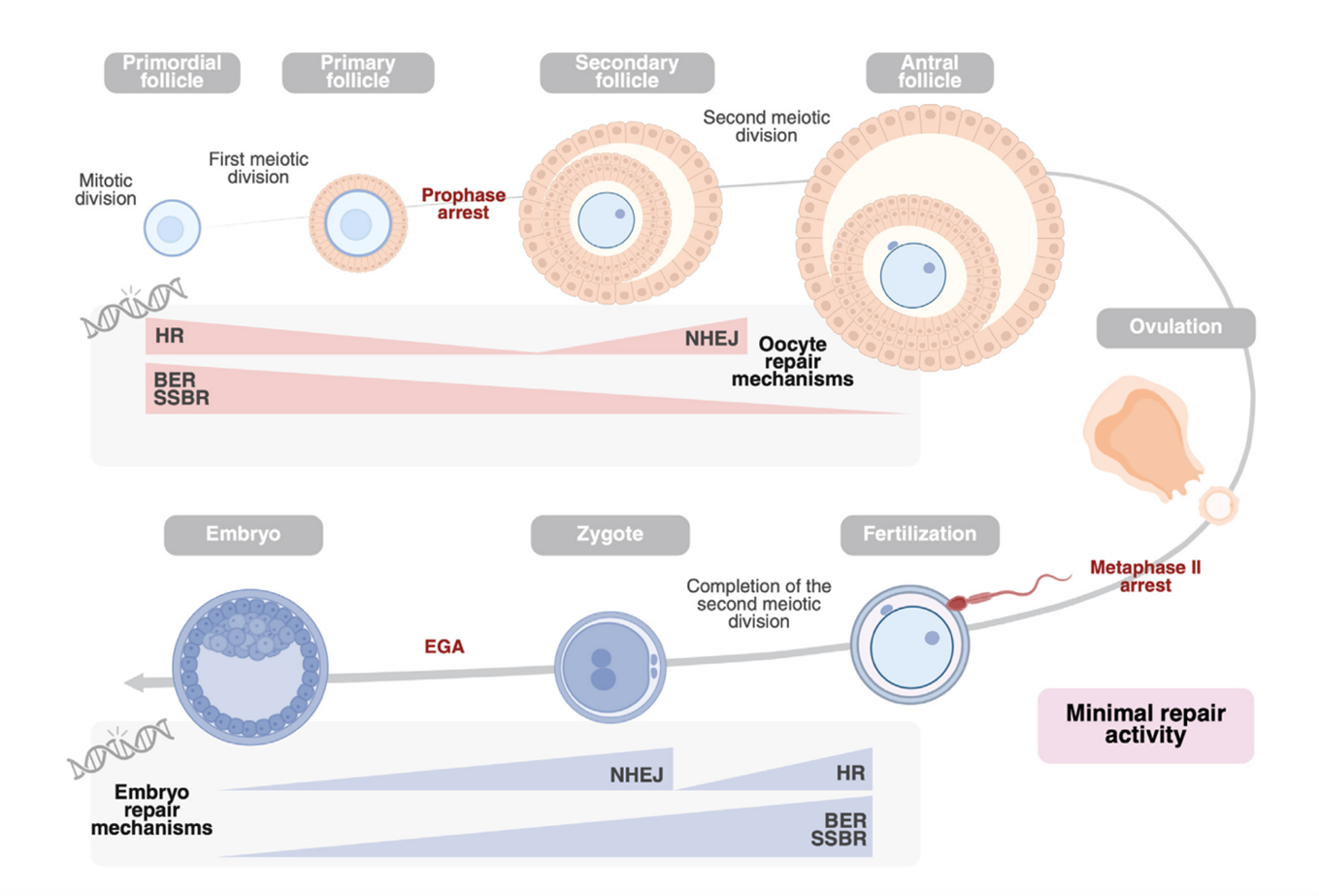
Quá trình sinh noãn là quá trình sinh học phức tạp và kéo dài. Bắt đầu từ việc các tế bào mầm nguyên thủy (Primordial Germ Cells - PGCs) di chuyển đến gờ sinh dục, tại đây chúng biệt hóa thành noãn nguyên bào.Các tế bào này sau đó bước vào giảm phân I nhưng tạm dừng ở kỳ đầu I (giai đoạn diplotene) dưới dạng các noãn bào sơ cấp (hay còn gọi là túi mầm, Germinal Vesicle - GV). Dưới tác động của đỉnh LH và các tín hiệu sinh học, noãn bào tái khởi động giảm phân I, hoàn tất việc phân chia để chuyển sang giảm phân II. Tuy nhiên, sự trưởng thành hoàn toàn chỉ đạt được khi có sự xâm nhập của tinh trùng, dẫn đến việc giải phóng thể cực thứ hai và hình thành hợp tử.
- Vai trò của vi môi trường và các tín hiệu điều hoà tại buồng trứng:
Vi môi trường buồng trứng (Ovarian Microenvironment) đóng vai trò không thể thiếu trong việc nuôi dưỡng và điều phối quá trình phát triển của noãn bào. Tế bào hạt (granulosa cells) bao quanh noãn bào cung cấp chất dinh dưỡng và các tín hiệu điều hòa thông qua mối nối khe (gap junctions), đồng thời tiết các yếu tố tăng trưởng như EGF (epidermal growth factor) có vai trò phá vỡ mối liên kết này khi cần tái khởi động giảm phân. Bên cạnh đó, con đường tín hiệu c-Kit/PI3K-PTEN-AKT đóng vai trò then chốt trong việc kích hoạt các nang noãn từ trạng thái nghỉ để phát triển qua các giai đoạn từ nang nguyên thủy đến nang trưởng thành. Sự trưởng thành cuối cùng của tế bào noãn là giai đoạn then chốt để đạt được khả năng thụ tinh. Thông qua ba cơ chế phối hợp và tái khởi động giảm phân I bằng cách can thiệp vào con đường tín hiệu cAMP/PKA, thúc đẩy bài tiết các yếu tố tăng trưởng EGF để phá vỡ mối nối khe giữa tế bào hạt và noãn, và điều hòa các hormone (như Follicular fluid meiosis activating sterol - FF-MAS) cùng tín hiệu canxi trung gian và đỉnh LH giúp tế bào noãn chuyển đổi từ trạng thái nghỉ sang kỳ giữa II (metaphase II - MII). Việc hoàn tất các thay đổi về nhân, bào tương và màng không chỉ đảm bảo sự tái tổ hợp nhiễm sắc thể chính xác mà còn chuẩn bị sẵn sàng cho tế bào noãn hoàn tất lần giảm phân thứ hai ngay khi có sự xâm nhập của tinh trùng [4].
- Vai trò của vi môi trường và các tín hiệu điều hoà tại buồng trứng:
Việc lựa chọn nguồn tế bào gốc đóng vai trò quyết định đến hiệu suất biệt hóa và đặc tính sinh học của tế bào đích. Hiện nay, các nghiên cứu về biệt hóa dòng noãn tập trung chủ yếu vào bốn nguồn tế bào gốc chính: Tế bào gốc phôi (Embryonic Stem Cells - ESCs), tế bào gốc đa năng cảm ứng (Induced Pluripotent Stem Cells - iPSCs), tế bào gốc dòng mầm và tế bào gốc sinh dục buồng trứng (Ovarian Stem Cells - OSCs).
Tế bào gốc phôi (Embryonic Stem Cells - ESCs): Biệt hóa từ khối tế bào bên trong (Inner Cell Mass - ICM) của phôi nang giai đoạn ngày 5 - 7. ESCs là tế bào gốc vạn năng thực thụ, có khả năng tự làm mới vô hạn và biệt hóa thành mọi dòng tế bào, bao gồm cả tế bào dòng mầm. Dù có tiềm năng lớn trong nghiên cứu tái tạo noãn, việc sử dụng ESCs người vẫn vấp phải rào cản đạo đức do phá hủy phôi và hạn chế trong tạo noãn tự thân.
Tế bào gốc đa năng cảm ứng (Induced Pluripotent Stem Cells - iPSCs): Được tạo ra bằng cách tái lập trình các tế bào soma (như nguyên bào sợi, tế bào máu) về trạng thái đa năng nhờ các yếu tố phiên mã (OCT4, SOX2, KLF4, c-MYC). Với đặc tính sinh học tương đương ESCs, iPSCs cho phép biệt hóa thành hPGCLCs (human Primordial Germ Cell-Like Cells) qua hệ thống nuôi cấy 3D với hiệu suất cao. iPSCs giải quyết các rào cản về vấn đề đạo đức liên quan đến phôi người và cho phép tạo ra dòng tế bào mang bộ gen của chính bệnh nhân, từ đó mở ra khả năng tạo noãn bào tự thân.
Tế bào gốc dòng mầm (Germline Stem Cells - GSCs / Female Germline Stem Cells - FGSCs): là các tế bào đơn năng được phân lập từ biểu mô bề mặt buồng trứng hoặc dịch nang noãn sau các chu kỳ chọc hút. FGSCs biểu hiện các dấu ấn dòng mầm đặc trưng và cung cấp nguồn tế bào không xâm lấn, dễ tiếp cận cho nghiên cứu.
Tế bào gốc buồng trứng (Ovarian Stem Cells - OSCs): Là quần thể tế bào gốc hiếm nằm trong biểu mô bề mặt buồng trứng trưởng thành, có khả năng tự làm mới và biệt hóa thành noãn bào mới ngay trong buồng trứng người lớn. Điểm đặc biệt của OSCs là chúng tồn tại tự nhiên trong vi môi trường buồng trứng, do đó có thể tương tác với các tế bào hạt và tế bào vỏ nang, tạo điều kiện biệt hoá thành noãn bào hiệu quả hơn khi đặt trong hệ thống nuôi cấy phù hợp và mở ra triển vọng lớn trong bảo tồn sinh sản cho bệnh nhân ung thư.
Tế bào mầm nguyên thủy dạng tế bào (Primordial Germ Cell-Like Cells - PGCLCs): Sản phẩm trung gian in vitro được cảm ứng từ ESCs hoặc iPSCs, mô phỏng giai đoạn tế bào mầm nguyên thủy (PGCs) trong phôi thai người. PGCLCs đóng vai trò quan trọng trong nghiên cứu cơ chế xác định dòng mầm người ở điều kiện có kiểm soát và là nguyên liệu cho các hệ thống nuôi cấy hướng đến noãn trưởng thành.
Dựa trên các nguồn tế bào gốc này, các nhà nghiên cứu đã xây dựng các mô hình biệt hoá noãn bào in vitro tương ứng. Mỗi mô hình khai thác đặc tính sinh học riêng của từng nguồn tế bào, tạo nên các chiến lược tiếp cận đa dạng trong nuôi cấy noãn bào in vitro.
a/ Mô hình tạo noãn từ tế bào gốc vạn năng (Pluripotent Stem Cells - PSCs)
Để tạo ra noãn trong ống nghiệm, các nhà khoa học sử dụng tế bào gốc để biệt hóa thành các tế bào giống tế bào mầm nguyên thuỷ (primordial germ cell-like cells - PGCLCs). Đây là bước đệm then chốt giúp nghiên cứu sự sinh sản của con người mà không cần can thiệp trực tiếp vào phôi thai tự nhiên.
Trên mô hình chuột: tế bào gốc phôi chuột (mouse Embryonic Stem Cells - mESC) được kích thích bằng activin và yếu tố tăng trưởng biểu bì (Epidermal Growth Factor - EGF) để tạo ra các tế bào giống biểu bì phôi (EPI like cells - EpiLC). Sau đó, dưới tác động của BMP4, LIF, SCF và EGF, mEpiLC biệt hóa thành mPGCLCs có đặc điểm tương tự tế bào mầm giai đoạn phôi E9.5.
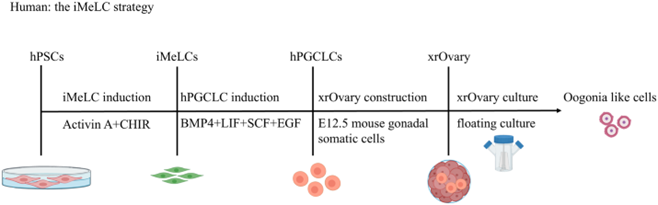
Trên mô hình người: Sử dụng hai chiến lược chính là iMeLC và 4i.
Chiến lược iMeLC (Incipient Mesoderm-Like Cell): Đây là quy trình cảm ứng hai giai đoạn. Giai đoạn đầu, PSCs được kích thích bởi Activin A và CHIR (chất cường vận tín hiệu WNT) để hình thành các tế bào giống trung bì sớm. Giai đoạn tiếp theo, các iMeLCs được nuôi cấy trong môi trường bổ sung BMP4, SCF, LIF và EGF để biệt hóa thành hPGCLCs.
Chiến lược 4i: Sử dụng bốn chất ức chế (CHIR99021, PD0325901, SB203580, SP600125) để duy trì PSCs ở trạng thái có khả năng biệt hóa dòng mầm cao, sau đó tiền cảm ứng với TGFβ/bFGF (hoặc Activin A) trước khi hình thành hPGCLCs bằng các yếu tố tăng trưởng.tế bào gốc được biệt hóa lại để mang đặc điểm giống hệt tế bào mầm trong cơ thể.
Sau đó, hPGCLC phân hóa thành tế bào giống noãn bào trong nuôi cấy tái tạo buồng trứng dị loài với tế bào soma buồng trứng phôi chuột. Những tế bào này bắt đầu biểu hiện các gen đặc trưng và thay đổi cấu trúc di truyền rất gần với noãn tự nhiên.
Ưu điểm của mô hình PSC là khả năng tự làm mới vô hạn và có thể tạo nguồn noãn tự thân (đặc biệt với iPSCs), đồng thời cung cấp hệ thống nghiên cứu cơ chế biệt hoá dòng mầm người mà không cần can thiệp vào phôi tự nhiên. Tuy nhiên, nhược điểm chính là quy trình tái lập trình phức tạp, hiệu suất thấp, nguy cơ biến đổi di truyền và ty thể (mtDNA), cũng như đòi hỏi môi trường tương tác phức tạp với tế bào sinh dưỡng tuyến sinh dục thai nhi để hoàn tất giảm phân. Về tính ứng dụng, đây là mô hình tiềm năng để nghiên cứu tạo noãn người và trong tương lai có thể cung cấp nguồn noãn tự thân cho bệnh nhân vô sinh mất hoàn toàn nguồn noãn, song vẫn cần thêm nhiều nghiên cứu trước khi ứng dụng lâm sàng. [4, 5, 6]
b/ Mô hình tạo noãn từ tế bào gốc đa năng cảm ứng (Induced Pluripotent Stem Cells - iPSCs)
iPSCs được tạo ra bằng cách tái lập trình tế bào soma trưởng thành (như nguyên bào sợi da, tế bào máu ngoại vi) thông qua việc đưa vào các yếu tố phiên mã Yamanaka (OCT4, SOX2, KLF4, c-MYC) bằng vector lentivirus, retrovirus, hoặc các phương pháp không tích hợp gen (episomal, mRNA, protein). Trên mô hình người, iPSCs được tiền cảm ứng thành iMeLCs bằng Activin A và CHIR99021, sau đó tiếp tục cảm ứng thành hPGCLCs (human Primordial Germ Cell - Like Cells) bằng BMP4, SCF, LIF và EGF trong hệ thống nuôi cấy 3D dạng cụm tế bào (aggregates). Từ hPGCLCs, các nhà nghiên cứu đã thiết lập hệ thống nuôi cấy tái tạo buồng trứng dị loài (Xenogeneic reconstituted ovary) bằng cách kết hợp hPGCLCs với tế bào soma buồng trứng chuột để tạo ra các tế bào giống noãn bào ở giai đoạn sớm, biểu hiện các dấu ấn đặc trưng như DAZL, SYCP3, ZP3 và có hình thái tương tự noãn nguyên bào người [4, 5, 7]. Những đổi mới gần đây bao gồm phương pháp chuyển đổi nhanh tế bào gốc đa năng cảm ứng người (hiPSCs) thành các tế bào giống noãn nguyên bào thông qua biểu hiện yếu tố phiên mã chỉ trong vài ngày, cũng như việc sử dụng các hệ thống đồng nuôi cấy với tế bào hỗ trợ buồng trứng, giúp cải thiện đáng kể tỷ lệ trưởng thành noãn in vitro (IVM) ở người so với môi trường nuôi cấy tiêu chuẩn. [6, 8]
Ưu điểm vượt trội của mô hình iPSC là hoàn toàn tránh được các vấn đề đạo đức liên quan đến phôi người, đồng thời cho phép tạo ra nguồn noãn tự thân từ chính tế bào soma của bệnh nhân, mở ra khả năng điều trị vô sinh cá thể hoá. iPSCs có thể được nhân rộng vô hạn trong nuôi cấy và về lý thuyết có thể cung cấp số lượng noãn bào không giới hạn. Nhược điểm chính bao gồm: nguy cơ đột biến gen và bất thường biểu sinh trong quá trình tái lập trình, nguy cơ hình thành khối u (teratoma) nếu còn sót tế bào chưa biệt hoá hoàn toàn, sự tích lũy bất thường ty thể (mtDNA) và hiệu suất biệt hoá thành noãn bào chức năng còn thấp.
Về tính ứng dụng lâm sàng, iPSC là mô hình đặc biệt phù hợp cho bệnh nhân đã mất hoàn toàn nguồn noãn do hoá trị hoặc xạ trị, bệnh nhân suy buồng trứng sớm (POI - Premature Ovarian Insufficiency), những người không thể hoặc không muốn dùng noãn hiến tặng. [9]
c/ Mô hình tạo noãn từ tế bào mầm nguyên thủy dạng tế bào (PGCLCs - Primordial Germ Cell-Like Cells)
PGCLCs (Primordial Germ Cell-Like Cells), tế bào mầm nguyên thủy dạng tế bào là sản phẩm trung gian then chốt trong toàn bộ lộ trình biệt hoá noãn bào in vitro từ PSCs hoặc iPSCs. Chúng đại diện cho giai đoạn tương đương với PGCs tự nhiên trong phôi thai người (khoảng ngày 12–16), trước khi di cư vào tuyến sinh dục và phát triển thành noãn nguyên bào. Hệ thống nuôi cấy 3D dạng cụm tế bào nổi (3D floating aggregate system) là phương pháp hiệu quả nhất để tạo PGCLCs từ PSCs/iPSCs người, được phát triển bởi nhóm nghiên cứu Saitou (Nhật Bản) và sau đó được tối ưu hoá rộng rãi. [3, 4, 6]
Quy trình nuôi cấy 3D tạo PGCLCs:
Giai đoạn 1: Tiền cảm ứng thành iMeLCs (incipient Mesoderm-Like Cells)
PSCs hoặc iPSCs được nuôi cấy đơn lớp (2D) trong môi trường bổ sung Activin A (50 ng/mL) và CHIR99021 (chất cường vận WNT, 3 μM) trong 36 - 48 giờ. Kết quả thu được quần thể iMeLCs, các tế bào giống trung bì sớm biểu hiện T (Brachyury) và EOMES cao, đóng vai trò là trạng thái tiền thân dễ cảm ứng dòng mầm nhất từ PSCs người. Đây là bước then chốt: bỏ qua giai đoạn này hoặc thay bằng EpiLC (như ở chuột) cho hiệu suất hPGCLC thấp hơn đáng kể ở người. [4, 9, 10]
Giai đoạn 2: Cảm ứng 3D thành hPGCLCs (Floating aggregates)
iMeLCs được phân tách thành tế bào đơn và tái tập hợp thành các cụm 3D (khoảng 3.000–5.000 tế bào/cụm) bằng cách nuôi cấy trong đĩa hoặc giếng hình chữ U có bề mặt không bám dính. Môi trường cảm ứng gồm BMP4 (500 ng/mL), SCF (100 ng/mL), LIF (1.000 U/mL) và EGF (50 ng/mL) trong nền GK15 (GMEM + 15% KSR) trong 4–6 ngày. Trong điều kiện này, các tế bào bên ngoài cụm phân hoá thành tế bào soma, trong khi một quần thể nhỏ tế bào bên trong cụm được bảo vệ khỏi tín hiệu BMP bởi chính các tế bào xung quanh, thu nhận đặc tính dòng mầm và trở thành hPGCLCs. Hiệu suất tạo hPGCLC từ hệ thống 3D có thể đạt 15 - 40% tổng số tế bào trong cụm, cao hơn đáng kể so với hệ thống 2D (dưới 5%). [3, 9, 10]
Giai đoạn 3: Xác định và phân lập hPGCLCs
hPGCLCs được nhận diện và phân lập bằng FACS dựa trên biểu hiện hai dấu ấn bề mặt đặc trưng là ITGA6 (integrin alpha-6, CD49f) dương tính và EPCAM (CD326) dương tính, kết hợp với PDPN dương tính và ITGB3 âm tính. Ở mức độ phân tử, hPGCLCs đặc trưng bởi sự biểu hiện cao các gen dòng mầm như BLIMP1/PRDM1, TFAP2C (AP2γ), NANOS3, PRDM14 và đặc trưng riêng ở người là SOX17 (thay vì SOX2 ở chuột) cùng với việc bất hoạt các gen đa năng OCT4/POU5F1, SOX2, NANOG. Bộ gen biểu hiện của hPGCLCs được xác nhận bằng RNA - seq có hồ sơ gần với PGCs người giai đoạn 4 - 7 tuần thai. [3, 9, 10]
Giai đoạn 4: Nuôi cấy tiếp trong buồng trứng tái cấu trúc (xenogeneic reconstituted ovary)
hPGCLCs được phân lập bằng FACS, sau đó phối hợp với tế bào soma buồng trứng phôi chuột (ngày phôi E12.5) theo tỷ lệ khoảng 1:10 để tái tạo thành cụm buồng trứng 3D. Cụm này được nuôi cấy trên màng lọc Transwell trong môi trường α-MEM bổ sung 10% FBS, SCF và BMP4 ở 37°C. Qua 3 - 4 tuần nuôi cấy, hPGCLCs tiến triển thành các tế bào giống noãn nguyên bào rồi đến noãn bào giai đoạn sớm, biểu hiện các dấu ấn đặc trưng của noãn bào người như DAZL, SYCP3 và ZP3. Mặc dù chưa đạt được noãn MII trưởng thành hoàn toàn in vitro ở người, hệ thống này đã chứng minh được tính liên tục của quá trình biệt hoá từ tế bào đa năng đến giai đoạn sớm của sinh noãn. [3, 4, 9, 10]
Hệ thống nuôi cấy 3D PGCLCs cung cấp một mô hình thực nghiệm có độ tin cậy cao và có kiểm soát để nghiên cứu cơ chế phân tử xác định dòng mầm người, cho phép theo dõi toàn bộ quá trình từ tế bào đa năng đến tiền thân noãn bào. Hệ thống 3D tạo môi trường vi cơ học gần hơn với in vivo, tế bào tạo ra có đặc điểm biểu hiện gen gần với PGCs tự nhiên. Tuy nhiên, hệ thống nuôi cấy này cũng tồn tại một số nhược điểm như chưa đạt được trưởng thành noãn bào MII hoàn toàn in vitro ở người, vẫn cần dựa vào tế bào soma buồng trứng chuột dị loài để hỗ trợ, hiệu suất tổng thể còn thấp và biến thiên giữa các dòng tế bào, nguy cơ bất thường biểu sinh tích lũy qua nhiều bước nuôi cấy.
Về tính ứng dụng, hệ thống PGCLC 3D hiện là nền tảng nghiên cứu quan trọng để hiểu cơ chế sinh noãn người và sàng lọc tác nhân ảnh hưởng sinh sản, đồng thời là bước đệm không thể thiếu trong các chiến lược tạo noãn in vitro trong tương lai. [3, 6, 9]
d/ Mô hình tạo noãn từ tế bào gốc dòng mầm (GSCs) và tế bào gốc dòng mầm cái (FGSCs)
Tế bào gốc dòng mầm (Germline Stem Cells - GSCs) là các tế bào có khả năng tự làm mới và biệt hoá thành giao tử, được chia thành hai nhóm theo giới tính: tế bào gốc tinh trùng (Spermatogonial Stem Cells - SSCs) ở nam và tế bào gốc dòng mầm cái (Female Germline Stem Cells - FGSCs, còn gọi là Oogonial Stem Cells - OSCs) ở nữ. FGSCs được phân lập lần đầu từ buồng trứng chuột trưởng thành bởi nhóm nghiên cứu của Tilly vào năm 2004 và sau đó từ buồng trứng người vào năm 2012, gây ra tranh luận khoa học lớn vì thách thức học thuyết kinh điển cho rằng phụ nữ không thể tạo thêm noãn sau khi sinh. [4] Về quy trình phân lập và nuôi cấy: FGSCs được phân lập từ biểu mô bề mặt buồng trứng (ovarian surface epithelium) bằng phương pháp phân tách miễn dịch từ tính (Magnetic-Activated Cell Sorting - MACS), phương pháp bám dính khác biệt hoặc phân tách dòng chảy huỳnh quang (Fluorescence Activated Cell Sorting - FACS) nhắm vào kháng thể bề mặt DDX4 (VASA). Sau khi phân lập, FGSCs có thể được duy trì trong nuôi cấy dài hạn trên lớp tế bào nuôi STO hoặc trong môi trường đặc biệt bổ sung GDNF, bFGF, LIF và EGF. Khi được chuyển vào hệ thống đồng nuôi cấy với tế bào hạt hoặc cấy ghép vào buồng trứng nhận, FGSCs có thể biệt hoá thành các noãn bào mới và hình thành nang noãn nguyên thủy. [2, 11, 12]
Bằng chứng thực nghiệm về tiềm năng của FGSCs trên mô hình động vật:
- Ở chuột, FGSCs được đánh dấu huỳnh quang và cấy ghép vào buồng trứng đã bất sản (bằng busulfan) đã tạo ra noãn bào mang nhãn hiệu của người cho, thụ tinh bình thường và sinh con khỏe mạnh.
- Ở người, hFGSCs được cấy vào mô vỏ buồng trứng người và ghép dị loài vào chuột thiếu hụt miễn dịch đã hình thành nang noãn người mới sau 1-2 tuần. Ngoài ra, các nghiên cứu còn xác định được quần thể FGSCs từ dịch nang noãn phế bỏ sau các chu kỳ chọc hút, cung cấp nguồn tế bào không xâm lấn và dễ tiếp cận cho nghiên cứu và ứng dụng. Về đặc điểm sinh học, FGSCs biểu hiện các dấu ấn dòng mầm như DDX4, DAZL, STRA8, MVH nhưng không biểu hiện các dấu ấn noãn trưởng thành như ZP1-3, GDF9, và có hoạt tính telomerase cao. [1, 2, 12]
Ưu điểm của mô hình GSC/FGSC là bản chất tế bào gốc dòng mầm đơn năng đã được xác định trước theo hướng tạo noãn, không đòi hỏi quá trình tái lập trình phức tạp, tránh nguy cơ hình thành khối u liên quan đến tế bào vạn năng. FGSCs cũng ít bị ảnh hưởng bởi các bất thường biểu sinh so với iPSCs. Nhược điểm là số lượng FGSCs trong buồng trứng rất nhỏ (ước tính dưới 0,01% tổng số tế bào buồng trứng), hiệu suất phân lập thấp và sự tồn tại của chúng ở người vẫn còn là chủ đề tranh luận trong cộng đồng khoa học. Hiện chưa có quy trình nuôi cấy chuẩn hoá hoàn toàn để biệt hoá FGSCs thành noãn bào trưởng thành có khả năng thụ tinh hoàn toàn in vitro ở người.
Về tính ứng dụng, FGSCs/GSCs là ứng cử viên hứa hẹn nhất cho tái tạo khả năng sinh sản ở phụ nữ trẻ mắc POI hoặc ung thư buồng trứng, đặc biệt trong các chiến lược cấy ghép nội vi (in vivo) hơn là nuôi cấy in vitro. [1, 2]
e/ Mô hình tạo noãn bào từ tế bào gốc buồng trứng (OSCs - Ovarian Stem Cells)
Sự tồn tại của tế bào gốc buồng trứng (OSCs) trong buồng trứng trưởng thành là một phát hiện đột phá, thách thức quan điểm cũ cho rằng phụ nữ sinh ra với một lượng trứng cố định và không thể tạo thêm. OSCs (tế bào gốc dòng mầm cái - female germline stem cells - FGSCs) là những tế bào hiếm, có khả năng phân chia và biệt hóa thành noãn bào mới, mở ra hy vọng điều trị vô sinh và suy buồng trứng. Dù còn nhiều tranh cãi về việc phân lập (do dễ nhầm lẫn với tế bào quanh mạch), các kỹ thuật tiên tiến như tách tế bào kích hoạt bằng huỳnh quang (Fluorescence-activated cell sorting - FACS) nhắm vào các dấu ấn bề mặt đặc hiệu (như DDX4) đã giúp thu được quần thể OSCs có độ tinh khiết và tỷ lệ sống cao.
Về đặc điểm sinh học, OSCs có hình thái tương tự tế bào gốc tinh trùng ở nam giới: nhân lớn, ít tế bào chất, hoạt tính telomerase cao và biểu hiện các gen đa tiềm năng nhưng chưa biểu hiện các dấu ấn của trứng trưởng thành. OSCs được tách chiết từ buồng trứng chuột hoặc người dựa trên kháng thể DDX4 thông qua phương pháp phân tách tế bào bằng từ tính (Magnetic-Activated Cell Sorting - MACS) hoặc phân tách tế bào bằng dòng chảy huỳnh quang (Fluorescence Activated Cell Sorting - FACS). Để kích thích sự phát triển, OSCs cần môi trường nuôi cấy đặc biệt hoặc hệ thống đồng nuôi cấy với các tế bào hỗ trợ.
Các nghiên cứu thực nghiệm đã chứng minh tiềm năng to lớn của loại tế bào này trên cả hai mô hình ở chuột và người:
- Ở chuột: OSCs sau khi tiêm vào buồng trứng nhận hoặc nuôi cấy trong cơ quan nhân tạo (organoids) đã tạo ra được trứng chức năng, thụ tinh thành công và sinh con khỏe mạnh.
- Ở người: Việc tiêm hOSCs vào mô vỏ buồng trứng và ghép dị loài đã cho thấy sự hình thành các nang noãn mới. Đặc biệt, các nhà khoa học đã tìm ra nguồn OSCs từ dịch nang noãn bị loại bỏ sau các ca IVF, giúp giải quyết bài toán thiếu hụt mô hiến tặng.
Việc xác nhận danh tính của các noãn bào nhân tạo này được thực hiện qua các dấu ấn sinh học chuẩn (như GDF9, ZP1-3).
Ưu điểm nổi bật của mô hình OSC là bản chất đơn năng đã được lập trình sẵn theo dòng mầm, không đòi hỏi quá trình tái lập trình phức tạp như PSCs, đồng thời tránh được các rủi ro về đạo đức liên quan đến sử dụng phôi người. OSCs từ mô vỏ buồng trứng trữ lạnh vẫn duy trì khả năng biệt hoá tương đương mẫu tươi, mở ra khả năng ứng dụng trong bảo tồn sinh sản. Nhược điểm chính là sự tranh cãi về sự tồn tại thực sự của OSCs ở người (do dễ nhầm với tế bào quanh mạch), hiệu suất phân lập thấp và chưa chứng minh hoàn toàn khả năng thụ tinh của noãn bào tạo ra.
Về tính ứng dụng, mô hình OSC đang trở thành nền tảng quan trọng trong việc tái tạo khả năng sinh sản cho phụ nữ suy buồng trứng sớm hoặc vô sinh do điều trị ung thư, đặc biệt có thể khai thác nguồn OSCs từ dịch nang noãn phế bỏ sau các chu kỳ chọc hút IVF. [2, 13]
Trong bối cảnh tỷ lệ vô sinh ngày càng gia tăng, y học tái tạo với trọng tâm là liệu pháp tế bào gốc đang mở ra những triển vọng đột phá trong việc khôi phục chức năng sinh sản ở nữ giới. Việc phát hiện và ứng dụng các tế bào gốc buồng trứng (OSCs) cùng tế bào gốc mầm (GSCs) được chứng minh là có khả năng kích thích sự tái tạo, duy trì chức năng dự trữ buồng trứng thông qua việc cải thiện số lượng nang noãn và điều hòa các nồng độ hormone như FSH, AMH và E2. [1] Đặc biệt, công nghệ này trở thành một giải pháp thay thế an toàn và đầy hứa hẹn cho những phụ nữ trẻ bị suy buồng trứng sớm (premature ovarian failure - POI) hoặc vô sinh do điều trị ung thư (CTRI), giúp tránh được các rủi ro từ việc kích thích estrogen hay nguy cơ tái đưa tế bào ác tính vào cơ thể khi thực hiện các chương trình bảo tồn khả năng sinh sản (fertility preservation - FP). [2]
Bên cạnh đó, các nguồn tế bào gốc khác như tế bào gốc trung mô dây rốn (umbilical cord MSCs - UCMSCs), tủy xương, dịch ối hay máu ngoại vi cũng cho thấy khả năng thúc đẩy sự phát triển nang noãn và biệt hóa thành các tế bào hỗ trợ như tế bào hạt. Đáng chú ý, các nghiên cứu đã xác nhận rằng OSCs phân lập từ mô vỏ buồng trứng trữ lạnh vẫn duy trì tỷ lệ sống cao và có khả năng biệt hóa thành các tế bào giống tế bào noãn tương đương với mẫu tươi. [2] So với tế bào gốc vạn năng (PSCs) vốn đòi hỏi quy trình tái lập trình phức tạp và đối mặt với rủi ro về di truyền ty thể (mitochondrial DNA - mtDNA), OSCs có lợi thế hơn do bản chất đơn năng và đã được lập trình sẵn theo dòng mầm, không yêu cầu thao tác di truyền sâu để thực hiện quá trình sinh noãn. [1, 4]
Tuy nhiên, việc chuyển đổi từ nghiên cứu thực nghiệm sang ứng dụng lâm sàng vẫn đối mặt với nhiều thách thức về mặt đạo đức và kỹ thuật. Trong khi OSCs có thể tạo ra tế bào trứng trưởng thành khi tương tác với các tế bào hạt tiền thân trong mô buồng trứng trưởng thành, thì các tế bào mầm nguồn gốc từ PSCs lại đòi hỏi môi trường tương tác phức tạp với tế bào sinh dưỡng tuyến sinh dục thai nhi để hoàn tất quá trình giảm phân [4]. Hơn nữa, dù đã có những thành công bước đầu trong việc tạo ra noãn bào in vitro, nhưng khả năng thụ tinh và tạo ra thế hệ sau khỏe mạnh từ các giao tử này vẫn cần được xác nhận thêm qua các nghiên cứu dài hạn. Do đó, việc hoàn thiện quy trình nuôi cấy, giải quyết nguồn cung cấp mô và tháo gỡ các rào cản đạo đức sẽ là chìa khóa để đưa liệu pháp tế bào gốc trở thành hiện thực trong điều trị vô sinh nữ. [1]
Nuôi cấy tạo noãn bào từ tế bào gốc buồng trứng là một lĩnh vực nghiên cứu đầy tiềm năng nhưng vẫn đang ở giai đoạn tiền lâm sàng. Các mô hình biệt hóa từ tế bào gốc vạn năng (PSCs) đã đạt được bước tiến quan trọng trong việc tạo ra các tế bào giống tế bào mầm tiền thân (PGCLCs) và noãn nguyên bào giai đoạn sớm. Song song đó, việc ứng dụng tế bào gốc buồng trứng (OSCs) với bản chất đơn năng dòng mầm đã chứng minh khả năng biệt hóa thành các tế bào giống tế bào trứng (OLCs) có tiềm năng thụ tinh, đặc biệt khi được nuôi cấy trong hệ thống đồng nuôi cấy với tế bào hạt hoặc buồng trứng tái cấu trúc. Ngoài ra, tế bào gốc trung mô (MSCs) từ tủy xương, dây rốn và nội mạc tử cung đóng vai trò hỗ trợ then chốt thông qua cơ chế điều hòa miễn dịch và tiết yếu tố tăng trưởng (growth factors) để phục hồi nồng độ hormone (FSH, E2) và tái tạo nang noãn. Các mô hình hiện nay đã cung cấp nhiều hiểu biết quan trọng về oogenesis in vitro, song chưa đủ bằng chứng để ứng dụng lâm sàng. Việc tiếp tục hoàn thiện mô hình nuôi cấy, kết hợp với các tiến bộ trong sinh học tế bào gốc trong tương lai sẽ quyết định vai trò của hướng tiếp cận này trong lĩnh vực hỗ trợ sinh sản.
- Wang J, Liu C, Fujino M, et al. Stem Cells as a Resource for Treatment of Infertility-related Diseases. Curr Mol Med. 2019;19(8):539-546. doi:10.2174/1566524019666190709172636
- Silvestris E, Minoia C, Guarini A, Opinto G, Negri A, Dellino M, Tinelli R, Cormio G, Paradiso AV, De Palma G. Ovarian Stem Cells (OSCs) from the Cryopreserved Ovarian Cortex: A Potential for Neo-Oogenesis in Women with Cancer-Treatment Related Infertility: A Case Report and a Review of Literature. Current Issues in Molecular Biology. 2022; 44(5):2309-2320. https://doi.org/10.3390/cimb44050157
- Aizawa E, Peters AHFM, Wutz A. In vitro gametogenesis: Towards competent oocytes: Limitations and future improvements for generating oocytes from pluripotent stem cells in culture. Bioessays. 2025;47(1):e2400106. doi:10.1002/bies.202400106
- Tian M, Zhang M. Advances in in vitro oocyte generation from pluripotent stem cells and ovarian stem cells. Front Endocrinol (Lausanne). 2025 Aug 11;16:1515253. doi: 10.3389/fendo.2025.1515253. PMID: 40862123; PMCID: PMC12375483.
- Yamashiro, C., Sasaki, K., Yokobayashi, S., Kojima, Y., & Saitou, M. (2020). Generation of human oogonia from induced pluripotent stem cells in culture. Nature Protocols, 15, 1560-1583. https://doi.org/10.1038/s41596-020-0297-5
- Smela M P, Kramme C, Fortuna P, et al., (2025). Rapid human oogonia-like cell specification via transcription factor-directed differentiation. EMBO Reports, 26, 1114 - 1143. https://doi.org/10.1038/s44319-025-00371-2
- Cheng H, Shang D, Zhou R. Germline stem cells in human. Signal Transduct Target Ther. 2022 Oct 2;7(1):345. doi: 10.1038/s41392-022-01197-3.
- Piechota S, Marchante M, Giovannini A, et al., (2023). Human-induced pluripotent stem cell-derived ovarian support cell co-culture improves oocyte maturation in vitro after abbreviated gonadotropin stimulation. Human Reproduction (Oxford, England), 38, 2456 - 2469. https://doi.org/10.1101/2023.03.27.534479
- Sasaki K, Yokobayashi S, Nakamura T, et al. Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells. Cell Stem Cell. 2015 Aug 6;17(2):178-94. doi: 10.1016/j.stem.2015.06.014. Epub 2015 Jul 16. PMID: 26189426.
- Murase Y., Yabuta Y., Ohta H. et al. Long‐term expansion with germline potential of human primordial germ cell‐like cells in vitro. EMBO J 39, EMBJ2020104929 (2020). https://doi.org/10.15252/embj.2020104929
- White Y, Woods D, Takai Y et al., (2012). Oocyte formation by mitotically-active germ cells purified from ovaries of reproductive age women. Nature medicine, 18, 413 - 421. https://doi.org/10.1038/nm.2669
- Woods, D., & Tilly, J. (2013). Isolation, characterization and propagation of mitotically active germ cells from adult mouse and human ovaries. Nature Protocols, 8, 966-988. https://doi.org/10.1038/nprot.2013.047
- Canosa S, Silvestris E, Carosso A, et al., (2025). Ovarian Stem Cells: Will the Dream of Neo-Folliculogenesis After Birth Become Real?. Obstetrical & Gynecological Survey, 80, 112 - 120. https://doi.org/10.1097/ogx.0000000000001360