CVPH. Ngô Thị Lan Phương
Đơn vị HTSS IVFMD FAMILY, Bệnh viện Đa khoa Gia Đình Đà Nẵng
Giới thiệu
Exosome là các túi nhỏ gồm hai lớp lipid, có đường kính từ 30 đến 150 nm, được tiết ra bởi nhiều loại tế bào và tạo điều kiện cho sự giao tiếp giữa các tế bào. Chúng bắt nguồn từ hệ thống nội bào và giải phóng vào môi trường ngoại bào. Các exosome mang các phân tử có hoạt tính sinh học, bao gồm axit nucleic, lipid và chủ yếu là protein, ảnh hưởng đến các tế bào đích và góp phần vào tương tác giữa các tế bào. Exosome đóng vai trò quan trọng trong cả chức năng sinh lý bình thường và các tình trạng bệnh lý, bao gồm các rối loạn sinh sản ở nam và nữ. Nhiều bộ phận khác nhau của đường sinh dục nam giải phóng exosome vào tinh dịch. Exosome tinh dịch, đặc biệt là mào tinh hoàn và tuyến tiền liệt, đã được chứng minh là ảnh hưởng đến khả năng sinh sản của nam giới. Hơn nữa, vai trò của exosome tinh dịch đã được chứng minh trong đường sinh sản của phụ nữ trong quá trình làm tổ và mang thai. Bằng chứng cho thấy lượng exosome trong tinh dịch khác nhau giữa tình trạng bình thường và bệnh lý, ảnh hưởng đến quá trình sinh sản. Do đó, exosome được coi là dấu ấn sinh học có giá trị không chỉ trong chẩn đoán mà còn có tiềm năng điều trị các tình trạng bất thường, đặc biệt là vô sinh [1].
Trong vô sinh nam, việc kiểm tra exosome tinh dịch có thể cung cấp thông tin chi tiết về chất lượng tinh trùng, chức năng sinh sản và các nguyên nhân tiềm ẩn gây vô sinh [2]. Hiện nay, exosome tinh dịch có tiềm năng đáng kể trong sinh sản, đóng vai trò là ứng cử viên cho dấu ấn sinh học để xác định bệnh nhân vô tinh và các dạng vô sinh nam khác [3]. Bài viết này nhằm mục đích cung cấp góc nhìn mới về exosome tinh dịch trong chẩn đoán và quản lý vô sinh nam.
Exosome: sinh học, thành phần và chức năng
-
Sự hình thành của Exosome
Các túi ngoại bào (Extracellular Vesicles-EV) gồm nhiều phân nhóm, trong đó có exosome, microvesicle và thể apoptosis, mỗi loại được phân biệt bởi nguồn gốc tế bào riêng biệt. Exosome có kích thước nhỏ hơn so với các EV khác. Ngoài sự khác biệt về kích thước, exosome còn khác nhau về mật độ, nguồn gốc tế bào, hình thái, thành phần phân tử và chức năng sinh học [4].
Các exosome có nguồn gốc từ nhiều loại tế bào khác nhau nhưng có chung các con đường sinh tổng hợp, tất cả đều bắt nguồn từ quá trình nội bào. Ban đầu, các túi nội bào được hình thành từ màng tế bào chất, sau đó trưởng thành thành các endosome sớm. Ở bước thứ hai, màng giới hạn của các endosome muộn trải qua quá trình nảy chồi, dẫn đến sự hình thành các intraluminal vesicles (ILV) bên trong tế bào. Các endosome muộn chứa các ILV này được gọi là các thể đa túi (multivesicular bodies-MVB). Cuối cùng, MVB có thể hợp nhất với lysosome để phân hủy hoặc hợp nhất với màng tế bào chất của tế bào thông qua các protein của bộ khung xương tế bào, dẫn đến việc giải phóng ILV thông qua quá trình xuất bào vào không gian ngoại bào và dịch cơ thể. Những ILV được giải phóng vào môi trường ngoại bào này được gọi là exosome [5]. Nhiều nghiên cứu đã xác định sự hiện diện của một loạt các protein thiết yếu cho quá trình sinh tổng hợp exosome. Quá trình phát triển này có thể diễn ra thông qua các cơ chế phụ thuộc phức hợp phân loại nội thể cần thiết cho vận chuyển (endosomal sorting complexes required for transport-ESCRT) hoặc thông qua các con đường độc lập với ESCRT [6].
-
Thành phần và chức năng sinh học của Exosome
Cấu trúc của exosome được đặc trưng bởi một màng nano hình cầu bao gồm một lớp kép lipid bao bọc chủ yếu là protein, lipid và nhiều loại axit nucleic, bao gồm DNA, mRNA, miRNA và các loại RNA không mã hóa khác [7]. Một nhóm protein thường được tìm thấy trong exosome là tetraspanin như CD9, CD63, CD81 và CD82, đóng vai trò là các dấu hiệu để nhận dạng exosome. Các protein này tham gia vào quá trình tổ chức màng, phân loại và sản xuất sản phẩm exosome, tạo điều kiện cho sự giao tiếp giữa các tế bào. Ngoài ra, exosome, có nguồn gốc từ khoang nội bào, chứa các protein vận chuyển và dung hợp màng như GTPase, flotillin, Annexin và protein họ Rab, tham gia vào quá trình lắp ráp, vận chuyển exosome và các sự kiện dung hợp. Các protein sốc nhiệt (Hsp70 và Hsp90) cũng là thành phần đáng chú ý của exosome, tham gia vào quá trình gấp protein, ổn định và phản ứng với căng thẳng của tế bào. Thành phần cụ thể của protein trong exosome có thể thay đổi tùy thuộc vào loại tế bào và điều kiện sinh lý [8]
[1]. Việc kết hợp các loại protein khác nhau này vào exosome trong quá trình sản xuất của chúng giúp tạo điều kiện thuận lợi cho tín hiệu giữa các tế bào và vận chuyển, góp phần vào vai trò chức năng của exosome trong quá trình giao tiếp giữa các tế bào và các quá trình sinh lý.
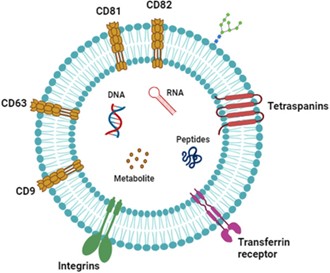
Hình 1. Các phân tử hoạt tính sinh học của exosome [1].
Exosome không chỉ chứa protein mà còn chứa nhiều loại lipid khác nhau, và thành phần lipid cụ thể của exosome có thể thay đổi tùy thuộc vào loại exosome. Thành phần lipid đa dạng của exosome đóng vai trò quan trọng trong việc hình thành cấu trúc, độ ổn định và chức năng của chúng [7].
Trong hệ thống sinh sản nam, một phân tích proteomic đã xác định được 1474 protein trong exosome tinh dịch của người. Phân tích bản thể gen cho thấy các protein exosome tinh dịch này chủ yếu liên quan đến exosome, tế bào chất và cytosol, tham gia vào các quá trình như con đường năng lượng, tăng trưởng tế bào và vận chuyển [9]. Đáng chú ý là mỗi đoạn của mào tinh tạo ra một quần thể epididymosome khác nhau, mỗi loại thể hiện thành phần lipid và protein riêng biệt. Các protein liên kết với mào tinh xác định hai nhóm túi riêng biệt: nhóm đầu tiên, giàu CD9 và các protein tetraspanin khác, tương tác với tinh trùng sống; nhóm thứ hai, được xác định bởi sự hiện diện của protein liên kết tinh trùng mào tinh 1 (ELSPBP1), tương tác với tinh trùng chết hoặc sắp chết [10]. Epididymosome vận chuyển một lượng lớn protein đến tinh trùng, bao gồm các nhóm enzyme, chaperone và protein cấu trúc quan trọng. Ngoài ra, epididymosome còn mang các protein bảo vệ chống lại sự tấn công của vi khuẩn và phân giải protein, và các enzyme glutathione peroxidase (GPX) giúp ngăn ngừa các phản ứng acrosome sớm và bảo vệ tinh trùng khỏi stress oxy hóa [11].
Prostasome là exosome chiếm ưu thế trong huyết tương tinh dịch. Các exosome này có thể được phân loại thành hai nhóm riêng biệt dựa trên kích thước và thành phần phân tử của chúng: các túi nhỏ hơn giàu protein liên quan đến sinh bệnh học thần kinh đệm 2 (glioma pathogenesis‐related-GLIPR2) và các túi lớn hơn giàu protein Annexin A1 (ANXA1) [12]. Các túi này chứa nhiều thành phần khác nhau, bao gồm enzyme, protein truyền tín hiệu, chaperone, protein vận chuyển và cấu trúc, protein liên kết GTP và các phân tử tín hiệu khác. Ngoài ra, chúng còn được làm giàu với hàm lượng cao phosphatase axit đặc hiệu tuyến tiền liệt (prostatic‐specific acid phosphatase-PAP), kháng nguyên đặc hiệu tuyến tiền liệt (prostate‐specific antigen-PSA), protease serine xuyên màng loại 2 (type 2 transmembrane serine protease-TMPRSS2), transglutaminase đặc hiệu tuyến tiền liệt (prostate‐specific transglutaminase-TGM4) và kháng nguyên tế bào gốc tuyến tiền liệt (prostate stem cell antigen-PSCA). Prostasome đã được chứng minh là có khả năng tạo ra adenosine triphosphate (ATP), tham gia vào quá trình chuyển hóa năng lượng [13].
Protein có trong prostasome thể hiện đặc tính chống oxy hóa, kháng khuẩn và kháng nấm. Đặc tính đông tụ của chúng có chức năng ngăn cản tinh trùng tiếp xúc với máu của phụ nữ, bảo vệ tinh trùng khỏi phản ứng miễn dịch trong đường sinh sản của phụ nữ bằng cách ức chế quá trình thực bào tinh trùng của bạch cầu đơn nhân và bạch cầu trung tính. Các protein vận chuyển trong prostasome được chuyển đến tinh trùng giống như mào tinh hoàn. Chúng đóng vai trò quan trọng trong sự sống còn và khả năng vận động của tinh trùng thông qua các con đường truyền tín hiệu phụ thuộc canxi. Thành phần lipid của prostasome được đặc trưng bởi sự chiếm ưu thế của các axit béo bão hòa hoặc không bão hòa đơn. Đáng chú ý, tính ổn định và độ cứng được tăng cường của màng prostasome có thể là do nồng độ cholesterol và sphingomyelin cao đáng kể [14].
Sự hiện diện của axit nucleic, bao gồm miRNA, mRNA và RNA nhỏ không mã hóa (sncRNA), trong exosome cho thấy rằng exosome có thể đóng vai trò là vật mang thông tin di truyền giữa các tế bào, cung cấp cơ chế giao tiếp giữa các tế bào và vận chuyển các phân tử điều hòa. Việc vận chuyển axit nucleic qua exosome có khả năng điều chỉnh biểu hiện gen và ảnh hưởng đến chức năng tế bào. Các kiểu biểu hiện miRNA thay đổi đã được quan sát thấy ở những bệnh nhân vô sinh. Kiểm tra RNA nhỏ (sRNA) đã chỉ ra rằng exosome có nguồn gốc từ tinh dịch giữ lại một cách chọn lọc nhiều loại sncRNA khác nhau, trong đó miRNA là phổ biến nhất, tiếp theo là YRNA, các đoạn RNA ribosome (rRNA), các đoạn tRNA (tRF), sncRNA có nguồn gốc từ gen mã hóa protein và piRNA, theo thứ tự giảm dần [15]. Hàm lượng RNA, đặc biệt là miRNA, của các exosome trong tinh dịch dường như thay đổi tùy thuộc vào tế bào gốc, cho phép nó phản ánh các điều kiện bệnh sinh lý của cơ quan ban đầu. Sự thay đổi này rất quan trọng khi xem xét các cấu hình biểu hiện miRNA trong các exosome như các dấu ấn sinh học có khả năng đáng tin cậy với ý nghĩa quan trọng đối với các ứng dụng chẩn đoán và điều trị. Các mào tinh hoàn đóng vai trò điều chỉnh biểu sinh của tinh trùng trong quá trình chúng di chuyển qua mào tinh hoàn. Các túi này có khả năng vận chuyển sRNA đến tinh trùng [16].
-
Exosome tinh dịch và khả năng sinh sản của nam giới
Exosome tinh dịch đóng vai trò quan trọng trong việc tạo điều kiện thuận lợi cho sự giao tiếp giữa huyết tương tinh dịch và tinh trùng, tác động tích cực đến các quá trình thiết yếu liên quan đến chức năng của tinh trùng. Chúng tham gia vào việc vận chuyển và phân phối các phân tử hoạt tính sinh học và các yếu tố tín hiệu khác nhau đến các tế bào tinh trùng đang phát triển. Các protein có trong exosome tinh dịch có nhiều chức năng khác nhau, tham gia vào sự tăng trưởng và duy trì tế bào, các con đường năng lượng và quá trình chuyển hóa protein. Chúng góp phần vào các quá trình tế bào như truyền tín hiệu, bám dính tế bào, vận chuyển màng và các hoạt động của enzyme. Nhiều nghiên cứu cho thấy các exosome có nguồn gốc từ đường sinh dục nam, bao gồm tế bào Sertoli, mào tinh hoàn và tuyến tiền liệt, đóng vai trò quan trọng trong quá trình phát triển tế bào mầm, quá trình trưởng thành của tinh trùng và ảnh hưởng đến kết quả thụ tinh [17].
.jpg)
Hình 2. Nguồn gốc và chức năng Exosome tinh dịch trong hệ thống sinh sản [1].
-
Exosome tinh dịch và quá trình sinh tinh
Ngoài vai trò về mặt cấu trúc, các nghiên cứu trên động vật đã chứng minh rằng các exosome có nguồn gốc từ tế bào Sertoli điều chỉnh sự tăng sinh, biệt hóa và apoptosis của tế bào mầm. Các túi này, được tiết ra thông qua cơ chế cận tiết, dễ dàng được các tế bào gốc tinh nguyên (spermatogonial stem cells-SSC) hấp thụ, cho phép giao tiếp động giữa các tế bào Sertoli và tế bào mầm - một tương tác cần thiết để duy trì hiệu quả sinh tinh [18]. Exosome có nguồn gốc từ tế bào Sertoli được làm giàu với các protein hoạt tính sinh học và axit nucleic điều chỉnh chức năng của SSC. Ngoài tế bào mầm, exosome tế bào Sertoli cũng ảnh hưởng đến sinh lý tế bào Leydig, tuy nhiên bằng chứng trên người còn hạn chế.
-
Exosome tinh dịch và sự trưởng thành của tinh trùng
Trong quá trình vận chuyển qua mào tinh hoàn, tinh trùng trải qua những biến đổi sinh lý và sinh hóa sâu sắc, bao gồm những thay đổi về hàm lượng protein, cholesterol, phospholipid và sncRNA. Những thay đổi này phần lớn được trung gian bởi các exosome mào tinh hoàn (epididymosome), cung cấp protein, RNA và lipid để bù đắp cho sự không hoạt động phiên mã của tinh trùng, do đó điều chỉnh sự tương tác giữa tinh trùng và tế bào [1]. Từ đó nhấn mạnh vai trò của chúng trong việc hình thành cấu hình RNA điều hòa của tinh trùng trong quá trình trưởng thành sau tinh hoàn.
Trong quá trình xuất tinh, tinh trùng gặp các exosome có nguồn gốc từ tuyến tiền liệt, hay prostasome, giàu cation hóa trị hai (Ca²⁺, Zn²⁺, Mg²⁺) và điều chỉnh nồng độ ion tinh trùng và khả năng vận động của roi thông qua cơ chế hợp nhất phụ thuộc vào pH. Sự chọn lọc prostasome có tính đặc hiệu cao, chủ yếu xảy ra ở đầu tinh trùng trong điều kiện pH trung tính và bicarbonate điển hình của đường sinh sản nữ. Gần phức hợp noãn-cumulus, prostasome có thể hợp nhất với các tế bào tinh trùng, cung cấp các thành phần trong tế bào chất và màng ảnh hưởng đến khả năng hoạt động, khả năng vận động và phản ứng acrosome [19].
Nhìn chung, những phát hiện này minh họa một chuỗi điều hòa liên tục do EV làm trung gian, bắt đầu từ các thể mào tinh hoàn (epididymosome) tái cấu trúc tinh trùng trong quá trình vận chuyển mào tinh hoàn, và mở rộng đến các thể tiền liệt tuyến (prostasome) tinh chỉnh chức năng tinh trùng trong quá trình xuất tinh và trưởng thành. Sự tích hợp này làm nổi bật vai trò trung tâm của các thể ngoại bào (exosome) như những chất mang năng động của protein, RNA và các phân tử tín hiệu, điều phối quá trình trưởng thành về mặt phân tử và chức năng của tinh trùng theo một cách thức phối hợp chặt chẽ và từng bước.
-
Exosome tinh dịch và khả năng hoạt động của tinh trùng
Trong quá trình trưởng thành sau tinh hoàn, tinh trùng trải qua những biến đổi lipid đáng kể và những thay đổi đáng chú ý trong môi trường proteome của chúng. Những thay đổi này rất quan trọng trong việc chuẩn bị tinh trùng cho quá trình trưởng thành khi chúng đi qua đường sinh sản của phụ nữ, nhằm mục đích tăng cường sự ổn định của màng sinh chất của tinh trùng, điều này rất cần thiết cho việc bảo quản tinh trùng lâu dài. Quá trình trưởng thành bao gồm một loạt các thay đổi về cấu trúc và chức năng được phối hợp, bao gồm các biến đổi trong thành phần màng tinh trùng, điều hòa hoạt động của enzym và quá trình phosphoryl hóa protein. Một dấu hiệu đặc trưng của quá trình trưởng thành là sự hoạt hóa quá mức của tinh trùng, đặc trưng bởi sự đập của roi không đối xứng và sự gia tăng Ca²⁺ trong tế bào chất. Để đảm bảo quá trình thụ tinh thành công, quá trình hoạt hóa phải được tính toán chính xác thời gian để xảy ra đồng bộ với quá trình rụng trứng. Sau khi xuất tinh, các thành phần của huyết tương tinh dịch - đặc biệt là các yếu tố mất hoạt hóa - bao phủ và ổn định bề mặt tinh trùng để ngăn ngừa hoạt hóa sớm. Prostasome bổ sung cho quá trình này bằng cách chuyển cholesterol và sphingomyelin đến màng tinh trùng, do đó duy trì sự ổn định và làm chậm phản ứng acrosome. Sự ức chế này phần lớn đạt được bằng cách duy trì tỷ lệ cholesterol trên phospholipid trong màng tinh trùng, vì hoạt hóa liên quan đến việc loại bỏ cholesterol khỏi màng [1].
Khi tinh trùng di chuyển qua đường sinh sản của phụ nữ, quá trình hoạt hóa sẽ khởi động một chuỗi các sự kiện truyền tín hiệu. Sự kích hoạt này làm tăng adenosine monophosphate vòng (cAMP) nội bào và thúc đẩy quá trình phosphoryl hóa tại các gốc serine, threonine và tyrosine. Exosome tinh dịch hiện được công nhận là yếu tố trung tâm trong quá trình điều hòa này, cung cấp các phân tử vận chuyển giúp tinh chỉnh các sự kiện liên quan đến hoạt hóa. Exosome huyết tương tinh dịch đóng vai trò trung gian ngăn ngừa tình trạng tinh trùng hóa sớm thông qua nhiều cơ chế khác nhau, chẳng hạn như ức chế dòng chảy cholesterol, giảm tính lưu động của màng đỉnh tinh trùng, điều hòa phản ứng acrosome và thu nhận các công cụ truyền tín hiệu Ca2+. Chúng điều chỉnh các kiểu phosphoryl hóa tyrosine, một sự kiện truyền tín hiệu nội bào quan trọng kiểm soát quá trình tinh trùng hóa.
-
Các thông số về exosome tinh dịch và tinh trùng
Chất lượng tinh dịch có thể bị ảnh hưởng bởi exosome của nam giới. Tác động của exosome lên tinh trùng đã được đánh giá bằng cách đánh giá khả năng vận động, trong đó khả năng hoạt động được xác định thông qua quá trình phosphoryl hóa tyrosine và phản ứng acrosome. Việc phân lập và mô tả đặc điểm của exosome tinh dịch từ những người đàn ông có mẫu tinh trùng bình thường, tinh trùng yếu và vô tinh sau cắt ống dẫn tinh cho thấy rằng các exosome thu được từ các nhóm khác nhau này biểu hiện các đặc điểm tương tự về hình dạng, kích thước, biểu hiện của các dấu hiệu và biểu hiện protein liên quan đến sự trưởng thành của tinh trùng và khả năng sinh sản. Exosome có nguồn gốc từ nam giới có tinh trùng bình thường đã được phát hiện có khả năng tăng cường khả năng vận động của tinh trùng và thúc đẩy khả năng sinh sản. Hiệu ứng này dường như được cho là do sự chuyển CRISP-1 từ exosome sang tinh trùng sau khi xuất tinh [2]. Proteomic của exosome phân lập từ huyết tương tinh dịch của nam giới bị normozoospermic và asthenozoospermic nặng cho thấy sự khác biệt về hàm lượng protein của chúng có khả năng ảnh hưởng đến đặc điểm tinh trùng. Điều này cho thấy sự khác biệt về chức năng tiềm ẩn giữa exosome lấy từ các cá thể tinh trùng bình thường và tinh trùng bất thường.
Ở nam giới bị giãn tĩnh mạch thừng tinh và vô sinh, sự rối loạn trong biểu hiện protein liên quan đến exosome đã được phát hiện. Người ta đã đề xuất rằng KIF5B và ANXA2 có thể được coi là những dấu ấn sinh học tiềm năng cho rối loạn chức năng exosome ở những bệnh nhân bị giãn tĩnh mạch thừng tinh. Ngoài ra, sự tích tụ angiotensinogen (AGT) trong huyết tương tinh dịch có thể dự đoán tình trạng suy giảm khả năng vận động của tinh trùng và mức độ phân mảnh DNA cao hơn ở tinh trùng từ bệnh nhân mắc chứng giãn tĩnh mạch thừng tinh một bên. Những phát hiện này nhấn mạnh mối liên hệ giữa biểu hiện protein huyết tương tinh dịch bất thường và sự gián đoạn cân bằng sinh lý có thể ảnh hưởng đến khả năng thụ tinh của tinh trùng, nhấn mạnh tầm quan trọng của việc hiểu rõ vai trò của các protein liên kết với exosome trong tình trạng vô sinh ở nam giới liên quan đến chứng giãn tĩnh mạch thừng tinh [20].
Exosome huyết tương tinh dịch có thể bảo vệ tinh trùng khỏi các tổn thương do lạnh. Việc thêm exosome tinh dịch người vào chất pha loãng tinh trùng đã cải thiện khả năng vận động của tinh trùng và tổng khả năng chống oxy hóa (total antioxidant capacit-TAC) so với việc chỉ sử dụng tinh dịch ban đầu. Xử lý tinh trùng bằng EV bao gồm exosome và microvesicle được phân lập từ các mẫu tinh dịch normozoospermic trước khi bảo quản đông lạnh cho thấy tác dụng bảo vệ tinh trùng. Exosome trong chất pha loãng đã cải thiện thời gian sống sót của tinh trùng và duy trì tính toàn vẹn của màng huyết tương của tinh trùng. Việc bảo toàn tính toàn vẹn của màng này được cho là do exosome liên kết trực tiếp với màng đầu tinh trùng, giúp tăng cường tính ổn định của cấu trúc màng. Ngoài ra, nghiên cứu đã chứng minh rằng exosome đóng vai trò tăng cường đặc tính chống oxy hóa của tinh trùng bằng cách duy trì tính toàn vẹn của màng tinh trùng [1].
-
Điều hóa quá trình thụ tinh bằng exosome tinh dịch
Trong đường sinh dục nữ, các tế bào tinh trùng trải qua một loạt các thay đổi sinh hóa và sinh lý đa dạng để có được các đặc tính thụ tinh và tiếp cận được noãn, nhấn mạnh động lực học phân tử phức tạp cần thiết cho quá trình thụ thai thành công. Khả năng của exosome tinh dịch trong việc vận chuyển các phân tử đến cả tế bào tinh trùng và tế bào miễn dịch trong đường sinh sản của nữ là điều khiến chúng trở nên linh hoạt trong sinh lý học tinh trùng. Khả năng thích nghi này cho phép exosome tác động đến thành phần phân tử và hành vi của tế bào tinh trùng, từ đó tăng cường khả năng thụ tinh của chúng. Hầu hết các protein tham gia vào quá trình này đều liên kết với bề mặt tinh trùng thông qua các thể mào tinh và prostasome. Chúng đóng vai trò quan trọng trong quá trình trưởng thành của tinh trùng và điều chỉnh tương tác giữa tinh trùng với đường sinh dục nữ, góp phần vào sự phát triển chức năng và sự sẵn sàng của tinh trùng để thụ tinh thành công [21].
-
Tác động của exosome tinh dịch lên nội mạc tử cung trong quá trình làm tổ
Tinh dịch, ngoài vai trò là môi trường vận chuyển tinh trùng để tạo điều kiện cho quá trình thụ tinh của noãn, còn ảnh hưởng đáng kể đến các mô sinh sản của phụ nữ. Nó gây ra những thay đổi về phân tử và tế bào giúp tối ưu hóa các điều kiện thụ thai và hỗ trợ việc thiết lập một thai kỳ thành công. Các thành phần hoạt tính sinh học trong tinh dịch hoạt động như các phân tử tín hiệu quan trọng, tạo điều kiện thuận lợi cho sự giao tiếp giữa tinh dịch và hệ thống sinh sản của phụ nữ [22]. Các EV được tìm thấy trong tinh dịch, chủ yếu là prostasome và mào tinh hoàn, được cho là có khả năng truyền tín hiệu do vai trò đã được xác lập của chúng trong khả năng sinh sản và điều hòa miễn dịch. Các exosome tinh dịch đã được chứng minh là có vai trò trong các phản ứng miễn dịch-viêm trong nội mạc tử cung. Chúng tham gia vào việc điều hòa vi môi trường tử cung liên quan đến sự làm tổ của phôi bằng cách điều chỉnh nồng độ chemokine và cytokine. Hơn nữa, exosome tinh dịch có thể kích thích tế bào gốc nội mạc tử cung tiết prolactin trong giai đoạn cửa sổ làm tổ [23].
Ngoài ra, thành phần của tinh dịch không chỉ ảnh hưởng đến khả năng sinh sản mà còn có tác động lâu dài đến sự phát triển của con cái. Mặc dù huyết tương tinh dịch không phải là yếu tố bắt buộc để sinh sản thành công, được chứng minh bằng phương pháp thụ tinh trong ống nghiệm thành công với các nguồn tinh trùng khác nhau, nhưng nó vẫn có thể ảnh hưởng đến sự phát triển của phôi và sức khỏe lâu dài của con cái [1].
-
Ứng dụng của exosome trong y học sinh sản
Protein tinh dịch đóng vai trò quan trọng trong quá trình phát triển tinh trùng, trưởng thành và thụ tinh. Trong khi đó, exosome tinh dịch chiếm khoảng 3% tổng lượng protein có trong huyết tương tinh dịch. Do đó, exosome tinh dịch và các thành phần protein của chúng, chẳng hạn như ANXA2, đóng vai trò quan trọng trong việc xác định chất lượng tinh trùng và khả năng thụ tinh. Các phương pháp điều trị vô sinh nam hiện nay bao gồm các phương pháp dựa trên tế bào như liệu pháp tế bào gốc trung mô hoặc các phương pháp điều trị không dùng tế bào như liệu pháp exosome. Exosome, nổi lên như một phương pháp điều trị mới trong việc giải quyết các tình trạng liên quan đến vô sinh nam, dường như đưa ra các giải pháp cho những thách thức gặp phải với liệu pháp MSC trong các điều kiện lâm sàng. Những tiến bộ gần đây trong các chiến lược dựa trên exosome mang đến một phương pháp tiếp cận không xâm lấn và đầy hứa hẹn để trẻ hóa quá trình sinh tinh và thúc đẩy tái tạo tinh trùng, có khả năng cách mạng hóa các phương pháp điều trị các rối loạn liên quan đến vô sinh nam. Tiềm năng điều trị của exosome có nguồn gốc từ AF trên mô hình chuột bị vô tinh đã chứng minh sự cải thiện đáng kể về cả chỉ số sinh tinh và các thông số tinh trùng sau khi tiêm exosome có nguồn gốc từ AF. Hơn nữa, một trong những thách thức là sự xuất hiện của các tổn thương tinh trùng do đông lạnh trong quá trình đông lạnh-rã đông và sự phá hủy tinh trùng do stress lạnh gây ra. Đáng chú ý, nghiên cứu đã xác định được tác dụng bảo vệ của exosome chống lại các tổn thương tinh trùng do stress lạnh, làm nổi bật tiềm năng của exosome trong việc giảm thiểu tổn thương do stress lạnh và tăng cường khả năng sống của tế bào tinh trùng [24].
Kết luận
Tóm lại, ngày càng có nhiều nghiên cứu nhấn mạnh vai trò then chốt của exosome tinh dịch không chỉ trong sinh sản nam mà còn ảnh hưởng đến sức khỏe của nữ giới. Tuy nhiên, vẫn còn nhiều câu hỏi chưa được giải đáp, hướng đến nhu cầu nghiên cứu trong tương lai để làm sáng tỏ và giải quyết những phức tạp xung quanh các cơ chế trung gian qua exosome trong sinh sản ở người. Tương lai của nghiên cứu về khả năng sinh sản ở nam giới đầy hứa hẹn, với tiềm năng phát triển các phương pháp tiên tiến để đánh giá sức khỏe sinh sản và triển khai các chiến lược nhằm cải thiện kết quả sinh sản. Việc tiếp tục nghiên cứu exosome, được hỗ trợ bởi các nghiên cứu tiền lâm sàng và các thử nghiệm lâm sàng được thiết kế chặt chẽ, là rất quan trọng để đánh giá toàn diện tính an toàn, hiệu quả và tác dụng lâu dài của các liệu pháp dựa trên exosome. Những tiến bộ này có tiềm năng cách mạng hóa lĩnh vực sinh sản ở nam giới, cung cấp các phương pháp điều trị tiên tiến và ít xâm lấn cho bệnh nhân.
Từ khóa: exosomes, fertility, reproduction, seminal fluid, spermatozoa
Tài liệu tham khảo
1. Mehdinejadiani, S., Azad, N., Dehghan, Z., Khosravizadeh, Z., Saberi, F., Rezaee, D., Pilehchi, T., Goudarzi, N., Salahi, E., & Mehdinejadiani, K. (2025). Role of Seminal Exosomes in Reproduction. Journal of Cellular Physiology, 240(11), e70106. https://doi.org/10.1002/jcp.70106
2. Murdica, V., Giacomini, E., Alteri, A., Bartolacci, A., Cermisoni, G. C., Zarovni, N., Papaleo, E., Montorsi, F., Salonia, A., Viganò, P., & Vago, R. (2019). Seminal plasma of men with severe asthenozoospermia contain exosomes that affect spermatozoa motility and capacitation. Fertility and Sterility, 111(5), 897-908.e2. https://doi.org/10.1016/j.fertnstert.2019.01.030
3. Candenas, L., & Chianese, R. (2020). Exosome Composition and Seminal Plasma Proteome: A Promising Source of Biomarkers of Male Infertility. International Journal of Molecular Sciences, 21(19), 7022. https://doi.org/10.3390/ijms21197022
4. Hessvik, N. P., & Llorente, A. (2017). Current knowledge on exosome biogenesis and release. Cellular and Molecular Life Sciences: CMLS, 75(2), 193–208. https://doi.org/10.1007/s00018-017-2595-9
5. Colombo, M., Raposo, G., & Théry, C. (2014). Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annual Review of Cell and Developmental Biology, 30, 255–289. https://doi.org/10.1146/annurev-cellbio-101512-122326
6. Colombo, M., Moita, C., van Niel, G., Kowal, J., Vigneron, J., Benaroch, P., Manel, N., Moita, L. F., Théry, C., & Raposo, G. (2013). Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. Journal of Cell Science, 126(Pt 24), 5553–5565. https://doi.org/10.1242/jcs.128868
7. Jeppesen, D. K., Fenix, A. M., Franklin, J. L., Higginbotham, J. N., Zhang, Q., Zimmerman, L. J., Liebler, D. C., Ping, J., Liu, Q., Evans, R., Fissell, W. H., Patton, J. G., Rome, L. H., Burnette, D. T., & Coffey, R. J. (2019). Reassessment of Exosome Composition. Cell, 177(2), 428-445.e18. https://doi.org/10.1016/j.cell.2019.02.029
8. Ha, D., Yang, N., & Nadithe, V. (2016). Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: Current perspectives and future challenges. Acta Pharmaceutica Sinica. B, 6(4), 287–296. https://doi.org/10.1016/j.apsb.2016.02.001
9. Yang, C., Guo, W. ‐b., Zhang, W. ‐s., Bian, J., Yang, J. ‐k., Zhou, Q. ‐z., Chen, M. ‐k., Peng, W., Qi, T., Wang, C. ‐y., & Liu, C. ‐d. (2017). Comprehensive proteomics analysis of exosomes derived from human seminal plasma. Andrology, 5(5), 1007–1015. https://doi.org/10.1111/andr.12412
10. D’Amours, O., Frenette, G., Bordeleau, L.-J., Allard, N., Leclerc, P., Blondin, P., & Sullivan, R. (2012). Epididymosomes transfer epididymal sperm binding protein 1 (ELSPBP1) to dead spermatozoa during epididymal transit in bovine. Biology of Reproduction, 87(4), 94. https://doi.org/10.1095/biolreprod.112.100990
11. Chabory, E., Damon, C., Lenoir, A., Kauselmann, G., Kern, H., Zevnik, B., Garrel, C., Saez, F., Cadet, R., Henry-Berger, J., Schoor, M., Gottwald, U., Habenicht, U., Drevet, J. R., & Vernet, P. (2009). Epididymis seleno-independent glutathione peroxidase 5 maintains sperm DNA integrity in mice. The Journal of Clinical Investigation, 119(7), 2074–2085. https://doi.org/10.1172/JCI38940
12. Aalberts, M., van Dissel-Emiliani, F. M. F., van Adrichem, N. P. H., van Wijnen, M., Wauben, M. H. M., Stout, T. A. E., & Stoorvogel, W. (2012). Identification of distinct populations of prostasomes that differentially express prostate stem cell antigen, annexin A1, and GLIPR2 in humans. Biology of Reproduction, 86(3), 82. https://doi.org/10.1095/biolreprod.111.095760
13. Ronquist, K. G., Ek, B., Morrell, J., Stavreus-Evers, A., Ström Holst, B., Humblot, P., Ronquist, G., & Larsson, A. (2013). Prostasomes from four different species are able to produce extracellular adenosine triphosphate (ATP). Biochimica Et Biophysica Acta, 1830(10), 4604–4610. https://doi.org/10.1016/j.bbagen.2013.05.019
14. Sullivan, R., & Saez, F. (2013). Epididymosomes, prostasomes, and liposomes: Their roles in mammalian male reproductive physiology. Reproduction (Cambridge, England), 146(1), R21-35. https://doi.org/10.1530/REP-13-0058
15. Vojtech, L., Woo, S., Hughes, S., Levy, C., Ballweber, L., Sauteraud, R. P., Strobl, J., Westerberg, K., Gottardo, R., Tewari, M., & Hladik, F. (2014). Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic Acids Research, 42(11), 7290–7304. https://doi.org/10.1093/nar/gku347
16. Reilly, J. N., McLaughlin, E. A., Stanger, S. J., Anderson, A. L., Hutcheon, K., Church, K., Mihalas, B. P., Tyagi, S., Holt, J. E., Eamens, A. L., & Nixon, B. (2016). Characterisation of mouse epididymosomes reveals a complex profile of microRNAs and a potential mechanism for modification of the sperm epigenome. Scientific Reports, 6, 31794. https://doi.org/10.1038/srep31794
17. Tian, H., Wang, X., Li, X., Song, W., Mi, J., & Zou, K. (2024). Regulation of spermatogonial stem cell differentiation by Sertoli cells-derived exosomes through paracrine and autocrine signaling. Journal of Cellular Physiology, 239(4), e31202. https://doi.org/10.1002/jcp.31202
18. Ruthig, V. A., & Lamb, D. J. (2022). Updates in Sertoli Cell-Mediated Signaling During Spermatogenesis and Advances in Restoring Sertoli Cell Function. Frontiers in Endocrinology, 13, 897196. https://doi.org/10.3389/fendo.2022.897196
19. Vickram, A. S., Samad, H. A., Latheef, S. K., Chakraborty, S., Dhama, K., Sridharan, T. B., Sundaram, T., & Gulothungan, G. (2020). Human prostasomes an extracellular vesicle - Biomarkers for male infertility and prostrate cancer: The journey from identification to current knowledge. International Journal of Biological Macromolecules, 146, 946–958. https://doi.org/10.1016/j.ijbiomac.2019.09.218
20. Panner Selvam, M. K., Agarwal, A., Sharma, R., Samanta, L., Gupta, S., Dias, T. R., & Martins, A. D. (2021). Protein Fingerprinting of Seminal Plasma Reveals Dysregulation of Exosome-Associated Proteins in Infertile Men with Unilateral Varicocele. The World Journal of Men’s Health, 39(2), 324–337. https://doi.org/10.5534/wjmh.180108
21. Samanta, L., Parida, R., Dias, T. R., & Agarwal, A. (2018). The enigmatic seminal plasma: A proteomics insight from ejaculation to fertilization. Reproductive Biology and Endocrinology: RB&E, 16(1), 41. https://doi.org/10.1186/s12958-018-0358-6
22. Schjenken, J. E., & Robertson, S. A. (2020). The Female Response to Seminal Fluid. Physiological Reviews, 100(3), 1077–1117. https://doi.org/10.1152/physrev.00013.2018
23. Rodriguez-Caro, H., Dragovic, R., Shen, M., Dombi, E., Mounce, G., Field, K., Meadows, J., Turner, K., Lunn, D., Child, T., Southcombe, J. H., & Granne, I. (2019). In vitro decidualisation of human endometrial stromal cells is enhanced by seminal fluid extracellular vesicles. Journal of Extracellular Vesicles, 8(1), 1565262. https://doi.org/10.1080/20013078.2019.1565262
24. Mahdavinezhad, F., Gilani, M. A. S., Gharaei, R., Ashrafnezhad, Z., Valipour, J., Nashtaei, M. S., & Amidi, F. (2022). Protective roles of seminal plasma exosomes and microvesicles during human sperm cryopreservation. Reproductive Biomedicine Online, 45(2), 341–353. https://doi.org/10.1016/j.rbmo.2022.03.033